One common misconception about evolution is that individual organisms evolve. It is true that natural selection acts on individuals: each organisms' traits affect its survival and reproductive success compared with other individuals.
But the evolutionary impact of natural selection is only apparent in the changes in a population of organisms over time.

For example, the medium ground finch (
Geospiza fortis), a seed-eater that lives on the Galapagos Islands.
In 1977, the population of birds was decimated by drought; of some 1,200 birds, only 180 survived. During the drought, small soft seeds were in short supply. Thus, the members of the population with larger and deeper beaks were better adapted to eat the more plentiful (but more difficult to open) large hard seeds. These birds were the ones that survived the drought and went on to reproduce, so that the next generation of
G. fortis had a larger beak than it had before the drought.
 |
The period of drought is marked by a dramatic change
in body and beak size & shape |
The finch population had evolved by natural selection.
However, it is important to understand that the individual finches did not evolve.
Each bird had a specific size beak which did not change during the drought. Rather, the proportion of large beaks in the population increased from generation to generation: the population evolved, not its members.
By focusing on evolutionary change in populations, we can define evolution on its smallest scales, called microevolution, as changes in allele frequencies in a population over generations.
Natural selection is not the only cause of microevolution. There are three main mechanisms that cause allele frequencies to change:
- Natural Selection
- Genetic Drift
- Gene Flow
Natural Selection
Individuals in a population exhibit variations in their heritable traits, and those with traits that are better suited to their environment tend to produce more offspring than those with traits that are not as well suited.
In genetic terms, the selection results in alleles being passed to the next generation in proportions that difer from those in the present generation.
For example, the fruit fly D. melanogaster has an allele that confers resistance to several insecticides, including DDT. This allele has a frequency of 0% in laboratory strains of fruit flies established from collected wild flies from the 1930s.

However, in strains established from flies collected after 1960, the allele frequency is 37%. It can be inferred that this allele arose by mutation between 1930 and 1960 as a response to the use of DDT as insecticide in farming.
As this example shows, an allele that confers insecticide resistance will increase in frequency in a population exposed to that insecticide. Such changes are not coincidental.
By consistently favoring some alleles over others, natural selection can cause adaptive evolution.
Genetic Drift

If you flip a coin 1,000 times, a result of 700 heads and 300 tails might make you suspicious about the coin.
But if you flip a coin only 10 times, and outcome of 7 heads and 3 tails would not be surprising. the smaller the number of coin flips, the more likely it is that chance alone will cause a deviation from the predicted result.
Chance events can also cause allele frequencies to fluctuate unpredictable from one generation to the next, especially in small populations - a process called genetic drift.
Figure 1 models how genetic drift might affect a small population of wildflowers. In this example, an allele is lost form the gene pool, but it is a matter of chance which allele is lost. Such unpredictable changes in allele frequencies can be caused by chance events associated with survival and reproduction.
 |
| figure 1 |
Perhaps a large animal, like a cow in a field grazing, stepped on the three C(w) C(w) individuals in generation 2, killing them and increasing the chance that only the C(r) allele would be passed to the next generation.
Allele frequencies can also be affected by chance events that occur during fertilization. for example, suppose two individuals of genotype C(r)C(w) had a small number of offspring. By chance alone, every egg and sperm pair that generated offspring could happen to have carried the C(r) allele and not the C(w) allele. Certain circumstances can result in genetic drift having a significant impact on a population. There are two different examples of this: the founder effect and the bottleneck effect.
1.) The Founder Effect
When a few individuals become isolated from a larger population, this smaller group may establish a new population whose gene pool differs from the source population; this is called the founder effect.
The founder effect might occur, for example, when a few members of a population are blown by a storm to a new island. Genetic drift, in which chance events alter allele frequencies, will occur in such a case if the storm indiscriminately transports some individuals, but not others, from the source population.
This effect is probably responsible for the high frequency of inherited disorders among isolate human populations.

For example, in 1814, only fifteen British colonists founded a settlement on Tristan da Cunha, a group of islands in the Atlantic Ocean between Africa and South America.
One of the colonists apparently was carrying a recessive allele for retinitis pigmentosa, a type of progressive blindness that only afflicts homozygous individuals.
Of the founding colonists' 240 descendants, four had the blindness. In the present day, the frequency of retinitis pigmentosa is ten times higher on Tristan da Cunha than in the rest of the population on Earth.
2.) The Bottleneck Effect

A sudden change in the environment, such as a fire or flood, may drastically reduce the size of a population. A severe drop in population size can cause the bottleneck effect, so named because the population has passed through a "bottle-neck" that reduces its size.
By chance alone, certain alleles may be over-represented among the survivors, others may be underrepresented, and some may be absent altogether.
Ongoing genetic drift is likely to have substantial effects on the gene pool until the population becomes large enough that chance events have less impact.
But even a population that passes through a bottleneck eventually recovers its size, although it may have low levels of genetic variation for a long period of time - a legacy of the genetic drift that occurred when the population was small.

For example, the African Cheetah almost went extinct due to hunting for its exotic pelt. Radical protections were put in place and the population recovered, but because there were so few surviving individuals to begin re-establishing the population that the allele variations among the current population is very low.
Thus, if one Cheetah is susceptible to a certain disease, most likely all other members of the species are as well. Bottlenecks can have a lasting impact on the survivability of a species based on which alleles survived the event.
The examples discussed highlight four key points:
- Genetic drift is significant in small populations
- Genetic drift can cause allele frequencies to change at random
- Genetic drift can lead to a loss of genetic variation within populations
- Genetic drift can cause harmful alleles to become fixed
Gene Flow
Gene flow is the transfer of alleles into or out of a population due to the movement of fertile individuals or their gametes.

For example, suppose a hypothetical group of wildflowers on the top of a hill, all red ( C(r)C(r)) homozygous.
At the bottom of the hill is a group of white wildflowers - C(w)C(w).
A bee flies up the hill with pollen from the white flowers and introduces the C(w) allele to the red population.
The introduction would modify the original population's gene frequencies in the next generation.
Because alleles are exchanged between populations, gene flow tends to reduce the genetic differences between populations.
If it is extensive enough, gene flow can in fact result in two populations combining into a single population with a common gene pool.
Alleles transferred by gene flow can also affect how well populations are adapted to local environmental conditions.

For example, researchers, studying the songbird
Parus major on the Dutch island of Vlieland observed survival differences between two populations on the island.
Females born in the eastern population survived twice as well as females born in the central population, regardless of where the females eventually settled and raised offspring.
This finding suggests that females born in the eastern population are better adapted than females born in the central population.
However, extensive field studies proved that the two populations are interconnected with high levels of gene flow (mating), so there should be little genetic difference between them.
Why then are eastern females better at surviving?
The answer lies in the unequal amounts of gene flow from the mainland. In a given year, 43% of the first-time breeders in the central population are immigrants from the mainland, compared with only 13% in the eastern population.
Birds with genotypes from the mainland apparently survive and reproduce poorly on Vlieland, and in the eastern population, selection reduces the frequency of these genotypes.
But in the central population, gene flow from the mainland is so high that it overwhelms the effects of selection.
As a result, females born in the central population have many immigrants genes, reducing their survivability.
Gene flow can also transfer alleles that cause adaptive prowess.

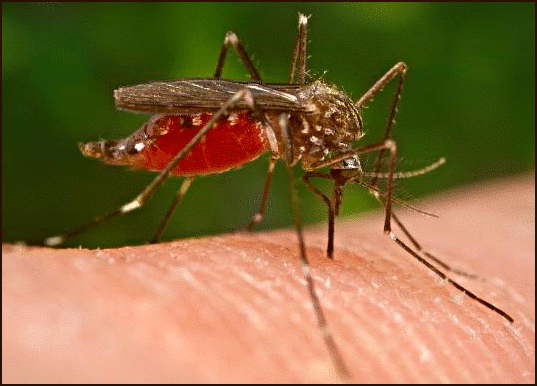
For example, gene flow has resulted in the worldwide spread of several insecticide-resistance alleles in the mosquito
Culex pipiens, a vector of West Nile virus and other diseases.
Each of these alleles has a unique genetic signature that allowed researchers to document that it arose by mutation in one or a few geographic locations. In their population of origin, these alleles increased because they provided insecticide resistance. These alleles were then transferred to new populations, where again, their frequencies increased as a result of natural selection.
Finally, gene flow has become an increasingly important agent of evolutionary change in human populations. Humans today move much more freely about the world that in the past. As a result, mating is more common between members of populations that previously had little to no contact, leading to an exchange of alleles and fewer genetic differences between those populations.
Concluding Thoughts
I have discussed the different forms of microevolution: natural selection, genetic drift, and gene flow. It has been shown how these forces cause evolutionary change that results in new species.
The next post will discuss why Natural selection is the only mechanism that consistently causes adaptive evolution.